|
|
 16-03-2009, 18:35
16-03-2009, 18:35
|
#1 |
|
Администратор

Регистрация: Nov 2006
Адрес: Израиль
Сообщений: 9,349
Поблагодарил: 713
Поблагодарили 6,059 раз(а) в 6,722 сообщениях Файловый архив: 54
Закачек: 12
|
Биохимия голодания (слабонервным не читать!) (Ха! Это вы еще цикл Кребса (он же цикл трикарбоновых кислот, он же дыхательный цикл) не видели) Прежде всего, хотелось бы отметить, что конкретные цифры могут сильно отличаться в разных исследованиях и источниках (до двух раз). Очевидно это зависит от участников, их веса, состояния здоровья, режима и т.д. Перефразируя Льва Толстого можно сказать, что все здоровые люди здоровы одинаково, а все больные – больны по своему. Существующие темы на форуме: Ацидотический криз Гликоген для голодания Биохимия http://www.xumuk.ru/biochem/124.html В. Свободная энергия гидролиза высокоэнергетических связей Предположительно в физиологических условиях энергия гидролиза АТФ до АДФ и неорганического фосфата равна примерно -50 кДж/моль. http://www.xumuk.ru/biochem/138.html А. Цитратный цикл Большая часть потребляемого в цитратном цикле ацетил-КоА получает ацетильные остатки, образовавшиеся в результате бета-окисления жирных кислот (т.е. 90% жира) (см. с. 166) и окислительного декарбоксилирования пирувата, катализируемого пируватдегидрогеназой (см. с. 136). Оба процесса протекают в матриксе митохондрий. http://www.xumuk.ru/biochem/140.html Цитратный цикл: функции клетки животных не в состоянии превращать жирные кислоты в глюкозу. Поэтому при голодании в организме прежде всего утилизируются не жиры, а белки. Высвободившиеся аминокислоты, напротив, могут превращаться и в жирные кислоты, и в глюкозу и, тем самым, поддерживать уровень сахара в крови (см. с. 300). (т.е. белки (читай мышцы) более универсальное топливо) Дополнительная информация В растениях и бактериях ацетил-КоА превращается в сукцинат в так называемом глиоксилатном цикле, тесно связанном с цитратным циклом. Эти организмы способны осуществлять анаплеротическую деградацию нейтральных жиров. В растениях глиоксилатный путь локализован в особых органеллах, глиоксисомах. (а вот бактерии и растения могут питаться жирами) http://www.xumuk.ru/biochem/148.html А. Аэробное и анаэробное окисление глюкозы В присутствии кислорода (в аэробных условиях) большинство клеток животных получают энергию за счет полного разрушения питательных веществ (липидов, аминокислот и углеводов), т. е. за счет окислительных процессов. В отсутствие кислорода (анаэробные условия) клетка может синтезировать АТФ (АТР) только за счет гликолитического разрушения глюкозы. Хотя такое разрушение глюкозы, заканчивающееся образованием лактата, дает незначительную энергию для синтеза АТФ, этот процесс имеет решающее значение для существования клеток при недостатке или в отсутствие кислорода. Жирные кислоты в виде ацилкарнитина попадают в матрикс митохондрий (см. с. 214), где подвергаются бета-окислению с образованием ацил-КоА (см. с. 166). Глюкоза в цитоплазме превращается в пируват путем гликолиза (см. с. 148). Пируват транспортируется в митохондриальный матрикс, где декарбоксилируется пируватдегидрогеназным комплексом (см. с. 136) с образованием ацетил-КоА. … Образующиеся из жирных кислот ацетильные остатки окисляются до CO2 в цитратном цикле (см. с. 138). Деградация аминокислот также приводит к ацетильным остаткам или продуктам, которые непосредственно включаются в цитратный цикл (см. с. 182). В отсутствие кислорода, т. е. в анаэробных условиях картина полностью меняется. … Поскольку энергия уже не может быть получена за счет деградации аминокислот, клетка становится полностью зависимой в энергетическом отношении от потребления глюкозы при гликолизе. … в клетках животных, функционирующих в анаэробных условиях, пируват восстанавливается до лактата, который поступает в кровь. … Продукция АТФ при этих процессах незначительна: при образовании лактата возникают только 2 молекулы АТФ на молекулу глюкозы. http://www.xumuk.ru/biochem/156.html Глюконеогенез Некоторые ткани, такие, как мозг и эритроциты, зависят от постоянного снабжения глюкозой. (это в «мирное время». На голоде мозг и нервы питаются и кетонами) Если получаемое с пищей количество углеводов недостаточно, необходимая концентрация глюкозы в крови может поддерживаться некоторое время за счет расщепления гликогена печенью (см. с. 158). Если истощены также и эти запасы, в печени запускается синтез глюкозы de novo, глюконеогенез (см. с. 302). Наряду с печенью высокой глюконеогенезной активностью обладают также клетки почечных канальцев (см. с. 320). Исходными соединениями в глюконеогенезе являются аминокислоты мышечной ткани. При длительном голодании это приводит к массивному распаду мышечного белка. Другими важными исходными веществами для синтеза глюкозы служат лактат, образующийся в эритроцитах и мышечной ткани при недостатке О2 (т.е. при интенсивных нагрузках), а также глицерин, образующийся при расщеплении жиров. Напротив, жирные кислоты не могут трансформироваться в глюкозу в организме животных, так как в данном случае деградация жирных кислот не является анаплеротическим процессом (см. с. 140). В организме человека за счет глюконеогенеза образуется несколько сотен граммов глюкозы в сутки. http://www.xumuk.ru/biochem/158.html Б. Баланс гликогена В организме человека может содержаться до 450 г гликогена, треть из которого накапливается в печени, а остальное — главным образом в мышцах. Содержание гликогена в других органах незначительно. Гликоген печени служит прежде всего для поддержания уровня глюкозы в крови в фазе пострезорбции (см. с. 300). Поэтому содержание гликогена в печени варьирует в широких пределах. При длительном голодании оно падает почти до нуля, после чего начинается снабжение организма глюкозой с помощью глюконеогенеза (см. с. 156). Гликоген мышц служит резервом энергии и не участвует в регуляции уровня глюкозы в крови. В мышцах отсутствует глюкозо-6-фосфатаза, поэтому гликоген мышц не может быть источником глюкозы в крови. (но при нагрузке он дает молочную кислоту, которая идет в дело в печени) По этой причине колебания содержания гликогена в мышцах меньше, чем в печени. http://www.xumuk.ru/biochem/164.html Деградация жирных кислот в печени Наиболее важным процессом деградации жирных кислот является бета-окисление (см. рис. 167) в митохондриях. При этом жирные кислоты вначале активируются в цитоплазме, присоединяясь к коферменту А [3]. Затем они с помощью транспортной системы (карнитинового челнока [4]; см. рис. 215) попадают в митохондриальный матрикс, где разрушаются в результате бета-окисления до ацетил-КоА. Образующиеся ацетильные остатки полностью окисляются до СО2 в цитратном цикле с освобождением энергии в виде АТФ (АТР).(при наличии углеводов или аминокислот. В последнем не уверен.) Если количество образовавшегося ацетил-КоА превосходит энергетическую потребность гепатоцитов, что наблюдается при высоком содержании жирных кислот в плазме крови (типичные случаи — голодание и сахарный диабет), то в гепатоцитах синтезируются кетоновые тела (см. рис. 305), снабжающие энергией уже другие ткани. http://www.xumuk.ru/biochem/166.html Деградация жирных кислот: бета-окисление А. Деградация жирных кислот: бета-окисление После попадания в клетки жирные кислоты активируются путем образования ацил-КоА Для этого нужны две богатые энергией ангидридные связи АТФ (см. с. 112). В матрикс митохондрий активированные жирные кислоты попадают в виде ацилкарнитина, который является трансмембранным переносчиком (см. с. 214). Деградация жирных кислот происходит в митохондриальном матриксе путем окислительного цикла реакций, при котором последовательно отщепляются С2-звенья в виде ацетил-КоА (активированной уксусной кислоты). Продуктами реакции являются ацетил-КоА и активированная жирная кислота, углеродная цепь которой короче на два углеродных атома по сравнению с длиной цепи исходной жирной кислоты. ……………………………………………………………………………………………………………………………… Для полной деградации длинноцепочечной жирной кислоты цикл должен многократно повторяться; например, для стеарил-КоА (18:0) необходимы восемь циклов. Образующийся ацетил-КоА может переноситься на оксалоацетат с образованием цитрата, промежуточного метаболита цитратного цикла (см. с. 140). При избытке ацетил-КоА в печени образуются кетоновые тела (см. с. 304).(жирные кислоты при наличии углеводов окисляются в митохондриях) Б. Энергетический баланс деградации жирных кислот Итак, при утилизации одной молекулы пальмитиновой кислоты синтезируются 106 молекул АТФ, что соответствует свободной энергии 3300 кДж/моль (106 х 30,5 кДж/моль АТФ). Выигрыш в энергии при деградации жирных кислот существенно выше по сравнению с распадом углеводов (32 молекулы АТФ на 1 молекулу глюкозы) и белков даже с учетом больших размеров молекул. Поэтому жиры представляют собой очень выгодную форму сохранения энергии. http://www.xumuk.ru/biochem/300.html Сердце и нервная ткань используют глюкозу в качестве источника энергии. Клетки сердечной мышцы являются в известном смысле "всеядными", так как они могут получать энергию и из других субстратов Б. Фаза пострезорбции При прекращении поступления пищи вскоре начинается фаза пострезорбции. Эта стадия начинается с изменения секреции гормонов поджелудочной железы: теперь А-клетки секретируют больше глюкагона, а В-клетки прекращают секрецию инсулина. Низкое соотношение инсулин/глюкагон в плазме крови запускает процесс промежуточного метаболизма в обратном направлении. Теперь организм должен вернуться к использованию собственных энергетических резервов. В организме начинается расщепление запасных веществ — гликогена, жиров, белков, и запускается производство макроэргических субстратов в печени. В печени происходит мобилизация гликогена (гликогенолиз, см. с. 158), Полученная глюкоза используется для обеспечения других тканей, прежде всего мозга, коры надпочечников и эритроцитов, не располагающих собственными резервами глюкозы. Если спустя несколько часов резервы глюкозы в печени окажутся исчерпанными, усиливается процесс глюконеогенеза (см. с. 156). Субстраты поступают из мышц (аминокислоты) и жировой ткани (глицерин). Высвободившиеся жирные кислоты используются печенью для синтеза кетоновых тел (кетогенез, см. с. 304), которые направляются в кровь и служат важнейшим источником энергии в пострезорбционной фазе. В мышцах разнообразные резервы глюкозы используются исключительно для собственных нужд (см. с. 238). Аминокислоты, образующиеся за счет медленного расщепления белков, поступают в печень и утилизируются в процессе глюконеогенеза. В жировой ткани гормоны инициируют липолиз с образованием глицерина и жирных кислот. Жирные кислоты служат источником энергии во многих тканях (за исключением мозга и эритроцитов). Важным приемником жирных кислот является печень, где они используются для синтеза кетоновых тел. (жир является источником двух видов сырья) 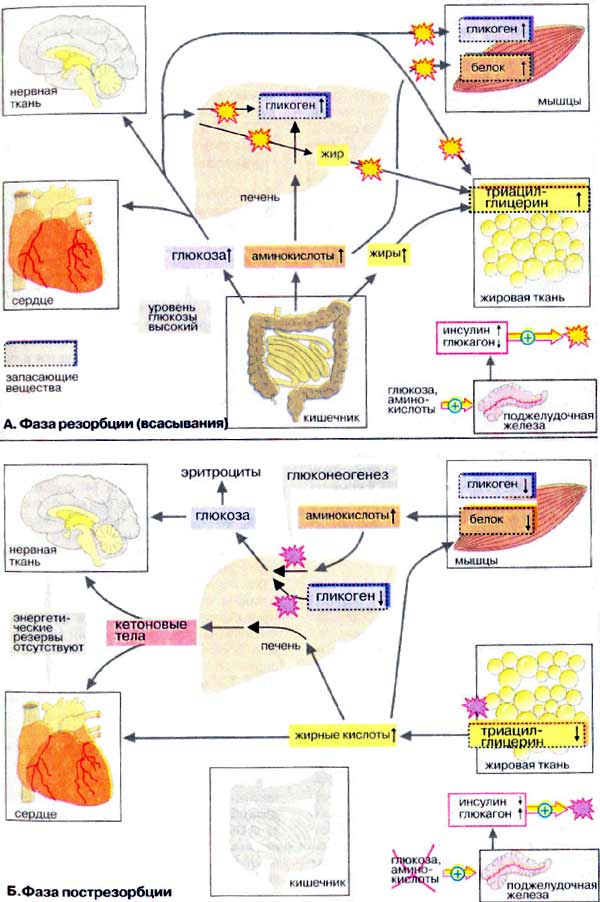 http://www.xumuk.ru/biochem/302.html А. Глюконеогенез: общие сведения Синтез глюкозы de novo (до 250 г в сутки (на голоде до 80 г.)) происходит в основном в печени (в мирной жизни). Процесс глюконеогенеза может идти и в почках, однако из-за небольших размеров почек их вклад в синтез глюкозы составляет всего 10%. (на голоде – 45%) Глюконеогенез контролируется гормонами. Кортизол, глюкагон и адреналин стимулируют этот процесс, а инсулин, напротив, подавляет. При глюконеогенезе в печени наиболее важными субстратами являются лактат, поступающий из мышечной ткани и эритроцитов, аминокислоты из желудочно-кишечного тракта (глюкогенные аминокислоты) и мышц (аланин), а также глицерин из жировых тканей. В почках в качестве субстрата служат главным образом аминокислоты (см. с. 320). Жирные кислоты и другие источники ацетил-КоА не могут использоваться в организме млекопитающих для биосинтеза глюкозы, поскольку ацетил-КоА, образующийся при бета-окислении в цитратном цикле (см. с. 140), полностью окисляется до СО2, в то время как в глюконеогенезе исходным продуктом является оксалоцетат. http://www.xumuk.ru/biochem/304.html А. Метаболизм липидов В фазе пострезорбции (см. с. 300), особенно в период поста или голодания, обмен липидов идет в обратном направлении, организм обращается к собственным запасам. В этих условиях жиры поступают из жировой ткани в кровь, переносятся в печень, распадаются в результате бета-окисления до ацетил-КоА и, наконец, превращаются в кетоновые тела. Б. Биосинтез кетоновых тел Кетоновые тела поступают из печени в кровь, где они хорошо растворимы. Концентрация кетоновых тел в крови возрастает в фазе пострезорбции (фаза голодания). Наряду с жирными кислотами 3-гидроксибутират и ацетоацетат в этот период являются основными энергоносителями. Ацетон, не имеющий метаболической ценности, удаляется через легкие. После 1-2 недели голодания кетоновые тела начинают использоваться в качестве источника энергии нервными тканями. Однако при этом для обеспечения цитратного цикла необходимо минимальное количество глюкозы. (жирные кислоты и кетоны используются как топливо, но для «растопки» нужна глюкоза) http://www.xumuk.ru/biochem/330.html А. Циклы Кори и аланина В клетках, не содержащих митохондрий (например, в эритроцитах), или в тканях при недостаточном снабжении кислородом (например, в активно работающих мышцах) АТФ (АТР) синтезируется за счет процесса превращений глюкозы в лактат, т. е. за счет процесса брожения (= анаэробного гликолиза, см. рис. 153). Лактат переносится кровью в печень, где в процессе глюконеогенеза с затратой АТФ (см. рис. 157) вновь конвертируется в глюкозу (цикл Кори). При интенсивной работе мышцы максимально активируется гликолиз. Продукт гликолиза, пировиноградная кислота (пируват) накапливается в цитоплазме и недостаточно быстро поступает в митохондрии, если они из-за недостатка кислорода не готовы к окислению пирувата. В анаэробных условиях пируват в реакции, катализируемой лактатдегидрогеназой (см. рис. 103) (заключительный этап гликолиза) восстанавливается до лактата. … Лактат диффундирует в кровь и поступает в печень, где конвертируется в глюкозу. Таким образом, образование лактата временно заменяет аэробный метаболизм глюкозы и частично переносит этот процесс из мышц в печень. Обратная связь, подобная циклу Кори, существует в цикле аланина, в котором также участвует пируват. Цикл аланина берет начало с протеолиза белков. Образующиеся аминокислоты в результате трансаминирования в присутствии ферментов превращаются в бета-кетокислоты (на схеме не приведено, см. рис. 181), которые в основном включаются в цикл трикарбоновых кислот (цитратный цикл) (см. рис. 183).Одновременно в реакции, катализируемой аланинтрансаминазой, аминогруппы из разных аминокислот переносятся на имеющийся субстрат, пируват. Образующийся аланин поступает в кровь и переносится в печень. Таким образом, цикл аланина служит каналом передачи азота и предшественников глюкозы в печень, которая является местом синтеза конечных продуктов азотистого обмена, например мочевины (см. рис. 185). Следует напомнить, что при анаэробном гликолизе образуются кислоты, которые, не принимая участие в последующем обмене, существуют в форме анионов. Поэтому при интенсивном анаэробном гликолизе рН мышечной клетки может понизиться настолько, что сокращение станет невозможным. Обычно этого не происходит благодаря быстрому выходу кислых метаболитов (лактата и пирувата) в кровь, которая также может оказаться закисленной (метаболический ацидоз) Б. Метаболизм белков и аминокислот Скелетные мышцы активно участвуют в метаболизме аминокислот. Это наиболее важный участок деградации разветвленных аминокислот (Val и Ile, см. с. 402). Ряд других аминокислот также деградируются преимущественно в мышцах. Одновременно идет ресинтез и высвобождение в кровь аланина и глутамина. Эти аминокислоты служат переносчиками азота, образующегося при расщеплении белков, в печень (цикл аланина) и почки (см. рис. 319). При голодании мышечные белки служат энергетическим резервом организма. Они гидролизуются до аминокислот, которые поступают в печень. Здесь углеродный скелет аминокислот конвертируется в промежуточные продукты цитратного цикла, в том числе в ацетоацетил-КоА и ацетил-КоА (см. рис. 183). Эти амфиболические соединения окисляются в цикле трикарбоновых кислот или включаются в процесс глюконеогенеза. (белки также являются топливом) Синтез и расщепление мышечных белков контролируются гормонами. Тестостерон и синтетические анаболики стимулируют биосинтез белка; напротив, кортизол подавляет образование мышечных белков. 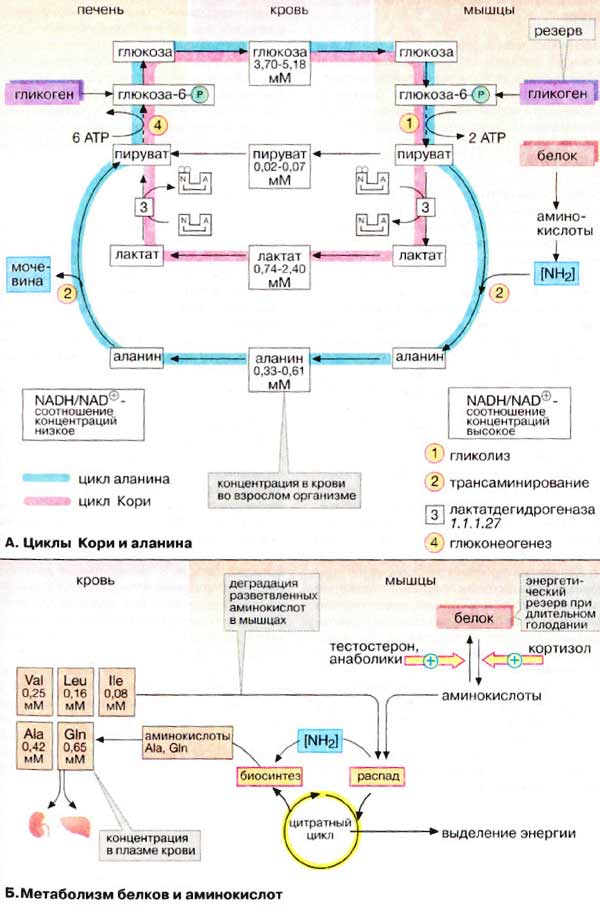 Последний раз редактировалось Vovec; 10-07-2010 в 17:03.. Причина: Ссылка на тему "Биохимия" |

|

|
| 21 пользователей сказали Vovec спасибо за это полезное сообщение: |
alagor (12-12-2009),
ASDF (16-03-2009),
Chudo-Yudo (13-07-2011),
inter (01-09-2009),
Ludmila_ (17-03-2009),
Maksenek (16-07-2012),
obelix (16-03-2009),
Olyy (31-03-2011),
Penelopa (19-03-2009),
pupsik (09-02-2011),
riskon (12-12-2009),
Stendal (20-04-2010),
Sytyj (27-02-2010),
Алена (16-03-2009),
Владомир (18-04-2017),
Елена_Ф (24-12-2009),
Мадам Фрекенбок (25-03-2009),
Семка (09-11-2009),
Слуцкий (08-06-2009),
троль (12-11-2009),
Эльвира (01-09-2009)
|
| Здесь присутствуют: 2 (пользователей: 0 , гостей: 2) | |
|
|
 Похожие темы
Похожие темы
|
||||
| Тема | Автор | Раздел | Ответов | Последнее сообщение |
| Биохимия | ROM | Теория и практика голодания, вопросы и советы | 214 | 11-08-2010 02:16 |
| Биохимия от Путина | Putin_IRL | Библиотека | 2 | 02-05-2007 03:36 |
| Спонсорские ссылки: | |
|
|
|






 Древовидный вид
Древовидный вид